-
SERT İSKELET YAPISINA SAHİP MERCANLARIN BAKTERİLER VE ÇÖZÜNMÜŞ ORGANİK MADDELER İLE BESLENMESİ
Yu. I. Sorokin
Sovyetler Birliği İç Denizler Biyoloji enstitüsü, Bilimler Akademisi,
Borok, Jaroslavl, USSR
ÖZET
Beslenme deneyleri Bismarck Archipolago resifinde bulunan 6 adet set iskelet yapısına sahip mercan türü üzerinde gerçekleştirilmiştir. Bu mercanların planktonik bakteriler ve çözülmüş organik maddelerle (protein hidrolizat) beslenme kabiliyetleri radyokarbon kullanılarak ispatlanmıştır. Hayvanlar tarafından hazmedilen etiketlenmiş yem olarak verilen organik karbon miktarı, polip bedeninin karbon hacminin %10-20’sine eşittir. Bazı planktonik alglerin mercanlar tarafından tüketilme ve hazmedilme oranı oldukça düşüktü. Mercanlar, planktonik bakterilerin hücrelerinde bağlı bulunan organik fosforu aynı oranda bulunan inorganik fosfattan daha aktif olarak kullanabilirler. Fosfor tüketim oranı günlük 3µg ‘dır.
Yonge ve Nicholl’un (1931) deneysel verilerine göre zooplankton sert iskelet yapısına sahip mercanların polipleri için temel gıda kaynağıdır. Ancak son zamanlardaki saha araştırmaları mercan resifleri üzerinden geçen zooplanktonların azaldığını göstermektedir (Johannes at al. 1970). Mercan polipleri kesinlikle simbiyotik zooxanthellaelerin ürettiği organik maddelerden faydalanır (Muscatine 1967; Franzisket 1969) fakat zooxanthellaeler tarafından gerçeklestirilen aktif fotosentez, tropikal sularda çok nadir olan geniş besin akışına ihtiyaç duyar. Poliplerin, partikül ve çözülmüş olmak üzere ilave besin kaynakları olduğuna inanılır.Stephens (1962) poliplerin 2-20 mg litre aralığındaki solusyon konsantresinden glikoz tüketebildiğini göstermiştir. Mercan polipleri genellikle, alg ve bakteri gibi planktonik organizmaları ayırabilecek kıllı uzantılara sahiptir. (Abe 1937; Roushdy and Hansen 1960). Mercanların planktonik bakterileri tüketmeleri ve sindirme yetenekleri Di Salvo (1969) ve Sorokin (1971) tarafından gösterilmiştir.
Resif oluşturan sert mercan poliplerinin olası beslenme mekanizmaları üzerine olan nicel çalışmalar, ki en zengin okyanuslarda bulunur - mercan resifi ekosistemlerinin enerji stoğunun hesaplanması için gereklidir (Odum ve Odum 1955). Batı tropikal Pasifik’e yaptığım seyahat sırasında çözülmüş organik maddelerin (dissolved organic matter: DOM) ve fosforun mercan polipleri tarafından sindirilmesini ölçme şansım oldu. Dr. R. Endean’a mercanların tanımlanması ve yapıcı eleştrilerinden ötürü minnettarım.
MALZEME VE YÖNTEMLER
Montipora, Pocillopora, Porites, Pavona ve Acropora’nın da dahil olduğu bazı ortak resif mercan türü örnekleri St. Andrew resifi ve Ninigo atoll (Bismarck Archipelago) adalarında da kurulan laboratuarlarda sürdürülen deneylerde kullanılmıştır. Mercanların dalları su altında makas ile kesilmiş ve kesilen kısım, sümüksü salgının salgılanmasını önlemek amacıyla, yumuşak plastik ile mühürlenmiştir. Bu şekilde hazırlanan dallar, kıyıya yakın yerde suya daldırılan bir akvaryumda bakıma alınmıştır. Deneyler simbiyotik zooxanhellaer tarafından olası işaretli CO2 emilimini önlemek amacıyla gece gerçekleştirilmiştir. Düşük miktarlarda CO2’nin değerlendirilmesi besin organizmalarının metabolizmasında yer almış olabilir.
Mercanların beslenme yoğunluğu radyokarbon yöntemi ile incelenmiştir. (Sorokin 1968, 1971a,b) Kültür olarak sağlanan doğal bakteriplankton, planktonik alg ve çözünmüş organik maddeler (DOM) etiketlenmiş ve besin olarak kullanılmıştır.
Bakterioplankton ince plankton ağı ile ön filtre yapılmış açık okyanustan alınan yüzey su örneğine (0.2-0.5 mg C litre) etiketli hidrolizat deniz alg proteini (Sorokin 1971) (radyoaktif) 14C eklenmesi ile etiketlenmiştir. 25-40 saat arasında 30 C’de karanlığa maruz kaldıktan sonra, etiketli hidrolizat neredeyse tamamen (%95-97) bakteriler tarafından tüketildi. Su HCI ile asitlendirdildi ve mikrobial metabolizmanın oluşturduğu etiketli CO2 hava kabarcıkarı ile sudan uzaklaştırıldı. Etiketli bakteriplankton içeren su, alkali solusyonu eklenerek PH 8.0 seviyesine getirildi. Sudaki çok küçük nanoplanktonik formasyon (oluşum) eksikliği membran filterelerinin doğrudan mikroskopileri tarafından gösterildi.
Planktonik alg, fotosentez sırasında etiketli 14C-COa2 sunumuyla gelişen kültürlerin içinde etiketlendi. Amphidinium klebsii ve yeşil flagellates türlerinin karışımı olan (L.Lanaskaya tarafından izole edilen ve korunan) kültürler ile birçok deniz canlısının filtre edilerek beslenebildiği etiketlenkmiş phytoplankton ve bakterioplankton en uygun düzeyde kullanıldı. (litre başına ıslak biyokütlenin 0.5-0.7 mg’si.)
Alg proteinin etiketli asit hidrolizatı, etiketli çözünmüş organik madde (DOM) olarak kullanıldı. (Sorokin 1971b). Hidrolizatın nötr çözeltisi steril ampuller içinde korundu. Bu çözelti kullanılmadan önce deniz suyunda 1/10 oranında seyretildi ve steril koşullarda bir membran filtreden geçirildi. (Filtre gözenek boyutu: 0.3µ). Deney kaplarının içindeki hidrolizat konsantrasyonu 0.3 mg C litre-1. Deneyler sırasında kapların içerisinde mikrop oluşumu ve bu nedenle partikül durumunda etiketli organik maddelerin besin kaynağı olarak (kullanılabilecek) DOM oluşumunu önlemek amacıyla, 50mg/litre oranında streptomycin eklendi. Kontrol deneyleri, bu yoğunluktaki streptomycin’in denizsuyu microflorası ile etiketli organik maddelerin mikrobik sindirimini 10 saat süre ile neredeyse tamamen durdurduğunu gösterdi. Aynı zamanda streptomycin’in varlığı mercanların ve planktonik canlılar da dahil olmak üzere diğer organizmaların hayatta kalma şansını arttırır.
Mercanların yoğunluğu ve beslenme verimliliği ölçütü olarak, sindirim değerleri Cα/C (alpha C/C) göstergesi kullanıldı. Bu gösterge sindirilen etiketli besin (24 saat başına hesaplanmış) içindeki karbonun, tüketici beden içindeki karbon miktarına olan yüzde oranı (Sorokin 1968). Bunu ölçmek için, yukarıda bahsedilen şekilde hazırlanmış 8-12 gr. mercan dalları, 1-2 litre ön filtreden geçirilmiş deniz suyu ile doldurulmuş akvaryum içine yerleştirildi. Bir porsiyon etiketli besin her akvaryuma ilave edildi. Bu şekilde 6-7 saatlik bırakıldıktan sonra, mercanlar 1 dakika süreyle deniz suyu ile yıkanarak yapışan etiketli besinden arındırıldı. Yıkanan mercanlar önceden HCO8 iyonlarından arındırmak için HCl ile kaynatılmış ve pH 8.2’ye ayarlanmış 200 ml deniz suyu dolu ölçeğe yerleştirildi. Deneyin sonunda mercan dokusu üzerindeki radyasyon (R) ve mercan tarafından deneyin ikinci aşamasında geliştirilen CO2 ölçüldü (ro).
Mercan dokularının alkalin hydrolizate R değeri ölçüldü. Bunlar, mercanların 3-5 ml gibi küçük oranlarda 0.1 N KOH solusyonunda kaynatılmasıyla hazırlandı. Dokular çözüldükten sonra artık madde basınçlı su ile mercan iskeletinden temizlenir. Hidrolizat HCl ile nötrlenen, hacmi 20ml’ye ayarlı ve belli parçaları metal kalıp levhaları üzerine dökülüp kurutuldu. Çözelti içindeki emilim, etiketsiz mercan dokusunun benzer çzöeltilerin içine katılan standart miktardaki etiketli alglerin azalmasının sayılması hesabı ile ölçüldü. ro değeri CO2 ’nin sudan damıtılması ve BaCO3 olarak çökelmesinden sonra tahmin edilir.
Eşzamanlı olarak, kontrol deneyleri, daha önce %10 formaldehyde ile sabitlenmiş mercanlar ile gerçekleştirildi. Kontrollerden elde edilen R değeri ve ro değeri ilgili deneysel veriden çıkartılır.
Etiketli besin miktarı deney sırasında mercanlar tarafından sindirilir. A, şu denklem kullanılarak hesaplanmıştır: A= Cr (R+ro) mg C koloni-1, Cr= karbon besinin ters özgül radyoaktivitesi, mg C.cpm-1. Sindirim index değeri (Ca/C = A(100)24%(Wt)-1Wt-1 ise W= deneyde kullanılan mercan kolonisi poliplerindeki organik karbon içeriği ve t= mercanın etiketli besin ile saat biriminde beslenme süresi. W’nin yaklaşık değerleri, polip gövdelerindeki ortalama karbon içeriğinin
mercanın ıslak ağırlığının %0.01’ine eşit olduğu varsayımına dayanılarak hesaplanmıştır. Bu oran literatürdeki temel veri (Odum ve Odum 1955) üzerinden ve Pocillopora polipleri değerleri ölçümleri üzerinden seçildi. Polipler güçlü formaldehyde solusyonu içinde sertleştirilip, ezilen mercanlardan izole edildi. Bunların karbon içeriği, ıslak yanma ile tespit edildi ve kolonide yer alan polip sayılarına göre hesaplandı.
Fosfatın mercanlar tarafından tüketilmesi 32P kullanılarak incelendi. İnorganik fosfat tüketim yoğunluğu ve partiküllü organik fosfat karşılaştırıldı. İki adet paralel deney yapıldı. Birinde mercanlar 6 saat boyunca streptomycin (500 meq ml-1) ve 7x105 cpm gerçek radyoaktivite ve ters özradyoaktiviteye (pr) 2x10-5 mg P.cpm-1 sahip inorganik etiketli fosfat içeren suda bekletildi. İkincisinde ise aynı miktarda etiketli fosfat, planktokik bakteri hücreleri içinde organik fosfat formunda eklendi. Bakteriplankton elde edebilmek için, inorganik fosfat payı ve 3mg glukoz, plankton ağından filtre edilmiş 1 litre deniz suyuna ilave edildi. İki gün karanlıkta bekletildikten sonra, etiketli inorganik fosfat neredeyse tamamen bakteriplankton tarafından tüketildi. 32P etiketli pakteriplankton içeren bu su, 1:3 oranında deniz suyu karıştırılarak seyreltildi ve deneylerde kullanıldı. Kontrollerde mercanlar Formalin kullanılarak sabitlendi.
Deneyin sonunda mercanlar dikkatlice yıkandı ve mercanların dokularında tüketilen 32P radyoaktivitesi, yukarıda bahsedildiği şekilde ölçüldü. Tüketilen 32P miktarı (D), D = R Pr µg P.colony-1 denklemi ile hesaplandı.
-
SONUÇLAR VE TARTIŞMA
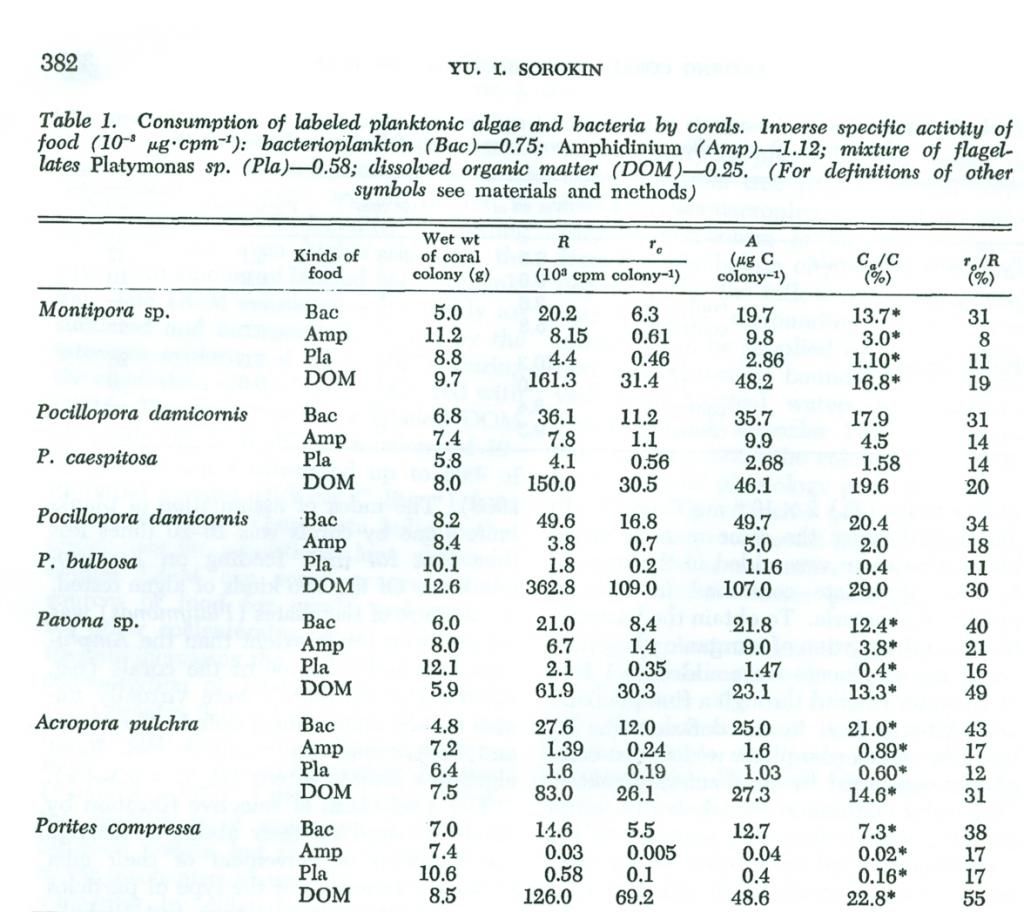
Mercanların değişik besinlerle beslenme yoğunluğu üzerindeki veri (Tablo1), üzerinde çalışılan tüm türlerin seçici birer filtre beslenicisi olduğunu gösterir. Mercanlar aktif olarak 1.2-1.6 x 106 cells ml-1 (ıslak biokütle 0.4-0.6 g m-3 ) yoğunluğunda planktonik bakteri maddelerini sindirmektedir. Bakterioplanktonun polipler tarafından sindirim endeksi (Ca/C), %10-20 değerlerine ulaşır. Bu da, küçük kabukluların etiketli besin ile beslenmelerinde bulunan seviye ile aynıdır (Sorokin 1968). Planktonik alglerin mercanlar tarafında sindirilme endeksi, onların bakterioplankton ile beslenme oranından 10-20 kat daha azdır. Test edilen 2 tür alg’de, Amphidinium örneklerinden daha az bir alanda Platymonas kullanıldı. Porites ve Acropra gibi bazı mercanlar neredeyse test edilen alg ile beslenememişlerdir. Montipora ve Pocillopora algi sınırlı bir oranda tüketip sindirmişlerdir (Ca/C=0.5-%4).
Mercanlar tarafından gerçekleştirilen bu seçici filtrasyon mekanizması, mercanların küçük kılımsı uzuvlarının (polip) hareket yönlerini, suda bulunan partiküllerin etkisine göre yönlendirebilme yetenekleri temeline dayanır (Yonge ve Nicholls 1931; Abc 1937). Yenilebilir partiküller varolduğunda, kılımsı uzuvlarının hareketi ağza doğru yönelir. Yenilebilir partiküller mevcut olmadığı durumlarda ise (sözgelimi carmine: bir çeşit kırmızı pigment), bir kimyasala olan reaktif etkisiyle (chemoreception) hareket tersine çevrilir ve böylece bu materyal polip gövde yüzeyinden uzaklaştırılır.
Kısa süren ikinci evre deney sırasında gelişen etiketli metabolik CO2 ‘nin göreceli miktarı, mercanlar, bakteri ile beslendiklerinde, alg ile beslendiklerinden daha fazlaydı. Bu da, tüketilen bakterinin alglerden daha fazla olduğunu gösterir. Mercanların bakterilerle beslenme oranı “ro : R” %30-40 idi ve bu da mercan poliplerinin yoğun metabolizmasına işaret eder.
Mercanların etiketli çözülmüş maddelerle beslenmesi ile ilgili deneyler, onların 0.2-0.5mg C Litre-1 gibi neritik (sığ deniz) sularındaki düşük moleküler ağırlıklı organik maddelerin doğal yoğunluğuna yakın düşük konsantrasyonlarda bu maddelerden faydalanma yeteneğini ortaya çıkartır. Mercanların DOM ile beslenme yoğunluğu, buna eşdeğer miktardaki etiketli bakterioplankton ile beslenmelerinden fazladır. Tüketilen DOM, daha önce DOM ile beslenmiş mercanların solunumu sırasında, etiketli CO2’nin yoğun dönüşümü tarafından gösterildiği gibi çabuçak sindirilmiş ve birleştirilmiştir. Etiketli DOM’un mercanlar tarafından tüketimi o kadar fazlaydı ki,10-12g ıslak ağırlıklı bir koloni 1 litre sudan kendi ağılığının%50’sine kadar (0.3mg C litre-1) tüketebilmişlerdir. Bu veri bize kesin koşullarda düşük moleküler ağırlıktaki DOM’un mercanlar için besin kaynaklarından biri olarak sunulabildiğini ve büyük olasılıkla diğer denizanası-gillerin saydam mukozalarının yüzey temasını arttırdığını göstermiştir. Kıyı sularındaki DOM konsantrasyonu litre başına 3-6mg C’ye denk gelmektedir ve organik maddenin önemli bir kısmı düşük moleküler ağırlıklı kesirler tarafından temsil edilir (Chailow 1971). DOM’un mercan polipleri tarafından tüketilme mekanizması hakkında az şey bilinmektedir, fakat bu işlem organik moleküllerin hayvanların bağırsaklarındaki mukozalarca sindirilmesi ile aynı olabilir (Ugolev 1967).
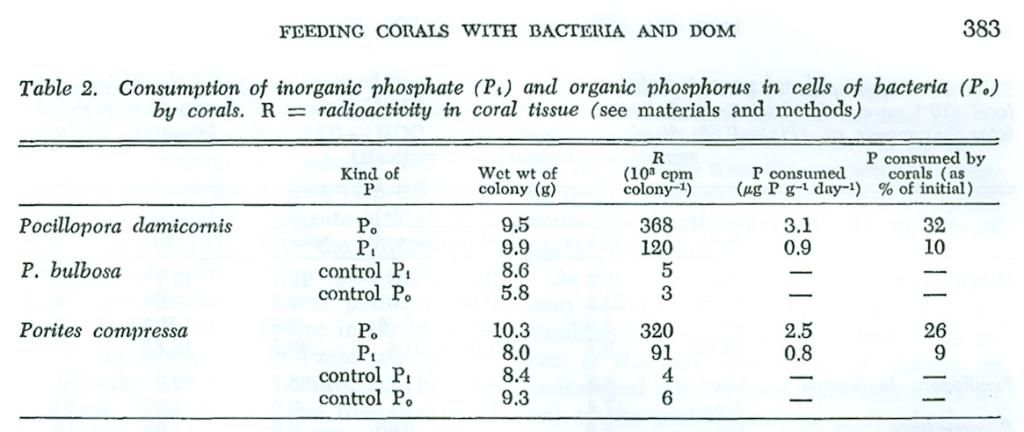
Etiketli inorganik ve organik fosfordan faydalanma üzerine yapılan deneylerin sonuçları (Tablo2), Pocillopora ve Porites gibi ortak mercanların bakterioplanktonların hücrelerinin özünde mevcut olan organik fosfor partiküllerini, aynı konsantrasyondaki inorganik fosfatdan daha fazla tükettiklerini göstermektedir. Görünen o ki, fosforun yanısıra, mercan polipleri ve onların simbiyotik zooxanthellaeleri bakteriyel beslenmenin sonucu olarak nitrojen, demir veya b12 vitamini gibi diğer önemli besinleri alabilmektedirler. Bu nedenle mercanların fitre beslenme kabiliyetleri, planktonik organizmaları yakalama kapasitelerinin yanısıra ve DOM’dan faydalanmaları sadece enerji ve poliplerce biosentez için organik maddeleri sağlamakla kalmaz aynı zamanda mercanların simbiyotik alglerinin fotosentez yapabilmeleri için besin de sağlar. Chlorophyll oluşumu ve fotosentez yoğunluğuna gelince, mercanlar coralin alg veya macrophytes gibi gerçek bitkilerden farklı değillerdir. Fakat onların ototropik (autotrophic: kendi kendini besleyen canlı) metabolizmaları esas olarak bitkilerin metabolizmasından farklıdır; zira bu, suyu çevreleyen mineral besinlerin akışına bağımlı değildir. Bu besinler tropikal sulara hakim organik olarak bağlı bulunan besinlerin tüketilmesiyle ve kısmen koloni içindeki besin döngüsüyle (Pomeroy ve Kuenzler 1969) sağlanabilir. Mercan fizyolojisinin bu özelliği belki mercanların mineral tuz(ları) bakımından zayıf tropikal sulardaki yüksek üretkenliğini sağlayan aşırı yoğun fotosentezlerini açıklayabilir.
Yalnızca gövde karbonu olarak alıkoyulan etiketli organik maddelerin sindirimi ölçüldü. Aslında bu, sadece denklemin bir tarafı olduğundan ve mercanlar tarafından organik maddelerin olası eşzmanlı salgılanması tespit edilmediğinden, bu deneyler net alınımı ölçmez. Pomeroy ve Kuenzley’e göre (1969), deneysel koşullar altında net fosfor salınımı yer alabilir. Bu nedenle, denge deneyleri mercanların komple nicel beslenme tanımı için gereklidir.
Çeviren : Ateş Hatipoğlu
REFERANSLAR
ABE, N. 1937. Feeding behaviour of Fungia nnd 15 other species of corals. Palau Trop. Sta. Stud. l: 429-450.
CHAILOW, K. M. 1971. Ecological metabolism in tho sea [in Russian]. Naukova dumkA, Kiev. 250 p.
D1 SALVO, L. H. 1969. Regeneration function and microbial ecology of coral reefs. Ph.D. thesis, Univ. Hawaii.
FRANZISKET, L. 1969. Riffcorallcn konnen Auto¬troph lebcn. Naturwisscnschaften 56: 144-146.
JOHANNES, R. E., S. L. COLES, AND N. T. KUENZEL. 1970. The role of zooplankton in the nutri¬ tion of some scloractinian corals. Limnol. Oceanogr. 15: 579-586.
MUSCATINE, L. 1967. Glycerol excretion by sym¬biotic algae from corals And Tridacna and its control by tho host. Science 156: 516--519.
ODUM, H. T., AND E. P. ODUM. 1955. Trophic
structure and productivity of a windward coral reef community on Eniwetok ntoll. Ecol. Monogr. 25:291-320.
PoMEROY, L. R., AND E. J. KVENZLEn. 1969. Phosphorous turnover by coral reef animals. Proc. 2nd Conf . Radioecology. AEC CONF- 670503, p. 474-482.
RousHDY, R. M ., AND V. K. HANSEN. 1960. Fil¬tration of phytoplankton by the octocoral Alcyonium digitatum. Nature 190: 649-650.
STEPHENS, G. C. 1962. Uptake of glucose from solution by the solitary coral Ftmgla. Science 131: 1532-1534.
SOROKIN, Yu. I. 1968. The use of C" in the studies of nutrition of aquatic animals.
Mitt. Int. Ver. Thcor. Angew. Lirnnol. 16. 41 p. --. 1971a. On the role of microflora in productivity of coral reef [in Russian]. J. Obshchei Biol. 32: 169-185.
--. 197lb. On the role of bacteria in pro¬ ductivity of tropical oceanic waters. Int. Rev. Gesamten Hydrobiol. 56: 1-48.
UGOLEV, A. M. 1967. Physiology and pathology of the by-wall digestion [in Russian]. Naukn, Leningrad.
YoNGE, 0. M., AND A. G. NICHOLLS. 1931. Stud¬ies on the physiology of corals. 4 and 5. Gt. Barrier Reef Expcd. 1928-29, Sci. Rep. 1: 135-176; 177-211.
Sunum: 14 Haziran 1972
Kabul : 23 Şubat 1973
Kimi birimler Türkçe karakterler tarafından temsil edilemediğinden ve ola ki orijinal dilinden makaleyi okumak isteyen olursa:
http://freepdfhosting.com/f2e55dc524.pdf
-
Merhaba ve mutlu yeni yıllar;
İki ayrı nano tankımda votka metodunu denedim ve faydasını gördüm. Dozlama olarak yabancı kaynaklarda excel tablosu şeklinde verilen tablo ile başladım.
Votka olarak Binboa kullandım. İlk denememde 9.haftada sonuçlarını aldım. 2.' sinde sonucu görmem daha kısa sürdü. Ancak yemlemenizi ve canlı yükünüzü adam akıllı kontrol altında tutmazsanız, dozlamayı bırakmanızın ardından nitrat tekrar yükselişe geçiyor. Fosfata olan etkisi daha düşük çünkü fosfatın kullanım oranı nitrata göre daha az...
Sürekli dozlamayı hiç denemedim ancak belirttiğim gibi yem ve canlı sayınızı abartmadığınız sürece kolay kolay tekrar yükselmiyor, yavaş bir grafik çiziyor.
Şu anda Fersan marka %5 asetik asit içeren beyaz sirke denemeye başladım.
Votka ve Sirke ile ilgili iki excel takip tablom var isteyenlere bağlantı atabilirim.
Konu mmoner tarafından (01-01-2015 Saat 11:26 AM ) değiştirilmiştir.

Yetkileriniz
- Konu Acma Yetkiniz Yok
- Cevap Yazma Yetkiniz Yok
- Eklenti Yükleme Yetkiniz Yok
- Mesajınızı Değiştirme Yetkiniz Yok
Forum Kuralları





 Alıntı ile Cevapla
Alıntı ile Cevapla